Prokariotinės chromosomos: naudingos pastabos apie prokariotines chromosomas
Štai jūsų pastabos apie prokariotines chromosomas!
Prokariotinės ląstelės genetinė informacija yra pernešama į nukleoplazmą ant bakterijų chromosomos. Jis susideda iš dvigubos sraigtinės DNR (deoksiribonukleino rūgšties) molekulės, niekada nesusijusi su baziniais baltymais, ir kai kuriose prokariotuose buvo parodyta, kad ji yra apvali.

Image Courtesy: hangthebankers.com/wp-content/chromosome.jpg
Todėl bakterinė chromosoma nėra struktūriškai homologinė su eukariotinės ląstelės branduolinėmis chromosomomis, o su organinių DNR, esančių eukariotinėse mitochondrijose ir chloroplastuose. Tikėtina, kad viena bakterinė chromosoma (viena labai ilga DNR molekulė) turi visą genetinę informaciją, būtiną prokariotinės ląstelės esminėms savybėms nustatyti.
Plazminiai vaistai: 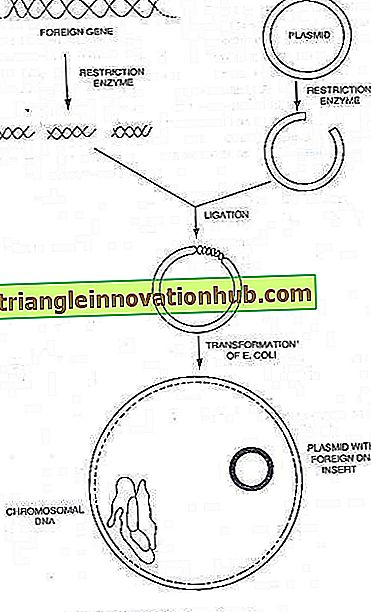

Daugelis bakterijų taip pat gali turėti mažas, papildomas chromosomų apykaitines DNR molekules, galinčias savarankiškai replikuotis, kurios yra žinomos kaip iki šiol ištirtos plazmidės. . DNR kiekis plazmidėje yra nuo 0, 1 iki 5 procentų bakterijų chromosomos.
Tai buvo aptikta Escherichia coli, kad yra du poravimosi tipai, o konjugacijos metu vienas partneris veikia tik kaip genetinis donoras arba vyras, o kitas tik kaip genetinis recipientas arba moteris. Kadangi vienintelė vyrų funkcija yra perduoti dalį savo DNR, ji neturi išlikti gyvybinga, o moteriškoji ląstelė turi likti gyvybinga, kad jame galėtų atsirasti zigotas.
Tai buvo nustatyta, kad bakterijų pažeidžiamumą lemia užkrečiamas genetinis elementas, kai vyriškos ir moteriškos bakterijos konjuguoja, kiekviena moterų ląstelė paverčiama vyru. Genetinis elementas, valdantis paveldėtą neturtingumo savybę, yra vadinamas F faktoriu (F = vaisingumas), jis perduodamas tik tiesioginiu ląstelių ryšiu. 1952 m. Lederbergas apibūdino terminą „plazmidė“ kaip genetinį pavadinimą visiems papildomiems chromosomų paveldimiems veiksniams, iš kurių F yra pavyzdys.
Dabar yra žinoma, kad bakterinės plazmidės yra mažos, apykaitinės DNR molekulės, kurios perneša genus savo replikacijai. Daugeliu atvejų jie taip pat turi genus, kurie suteikia šeimininkui ląstelių naujas savybes, pvz., Atsparumą vaistams arba toksinų gamybą. Daugelis plazmidžių turi genus, kurie reguliuoja konjugacijos procesą. Taigi, konjugacija yra mechanizmas, kuris bakterijų ląstelei yra priskirtas plazmidės, kurios normalus rezultatas yra plazmidės DNR perdavimas.
Plazminių tipai:
Dauguma plazmidžių buvo klasifikuojami pagal šeimininko savybes. Taigi yra gram-neigiamų bakterijų, Staphylococcus aureus penicilinazės plazmų, Pseudomonas degraduojančių plazmidžių ir kt. Faktorių (R = atsparumas) ir COI faktoriai (COI = kolicinogenai).
Plazminių savybės:
Visų žinomų plazmidžių struktūrą sudaro apvalios, dvigubos DNR molekulės. Keletas jų turi molekulinį svorį nuo 5 × I 7 iki 7 × 10 7 . Vienas (R faktorius) molekulinė masė yra tik 1 × 10 7, o kai kurie paslaptingi plazmidai yra dar mažesni. Kadangi DNR kiekis, reikalingas norint koduoti vidutinį polipeptidą, kurio molekulinė masė yra 40 000, yra apie 6 × 107, F1 ir kiti panašaus dydžio plazmidai gali turėti iki 100 genų.
Genų išraiška prokariotuose:
Bakterinės chromosomos:
Dabar nustatyta, kad bakterijų ląstelių centre yra bakterinė chromosoma, kuri nėra pajėgi mitozinėms ir meiotinėms dalims ir nėra uždaroma į tam tikrą branduolinę membraną. Vidutinėje bakterinėje ląstelėje yra vienas tūkstantis tipiškos eukariotinės ląstelės DNR kiekis.
Bakterinė chromosoma yra plonas, pailgos lanksčios ir apykaitinės DNR molekulės gijos, susietas su nedaugeliu baltymų. Bakterinės ląstelės gali labai greitai suskaidyti. Ląstelių pasidalijimas baigiamas padvigubinant visas ląstelių sudedamąsias dalis, o po to ląstelės padalijamos į dvi dukterines ląsteles. Pirmasis ląstelių pasidalijimo žingsnis yra DNR dubliavimas.
Genetinė rekombinacija bakterijose:
Tai yra procesas, kuriame genetinės medžiagos, esančios dviejuose atskiruose genomuose, sujungiamos į vieną vienetą. Bakterijose rekombinacija vyksta (1) transformacija, (2) transdukcija ir (3) konjugacija. Šie reiškiniai jau buvo išsamiai aptarti ankstesnėse dalyse, antraštėje „seksualinė reprodukcija bakterijose“.

Indukcija ir represijos:
Viruso genetinis potencialas yra ribotas ir jo replikacijai jis iš esmės naudoja šeimininkus. Kita vertus, bakterijos gali sintezuoti savo sudedamąsias dalis iš paprastų druskų ir cukraus. Bakterija, Escherichia coli, turi apie 2500 genų. Bakterija išlaiko savo vidaus ekonomiką tik leisdama vienu metu veikti keliems genams.
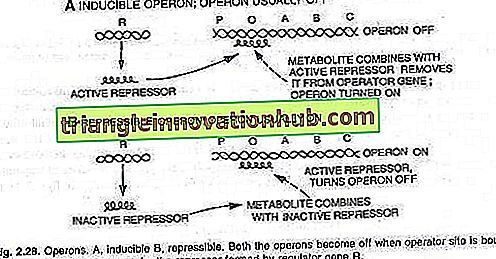
Tai reiškia, kad genai bus įjungti arba išjungti, kaip reikalaujama. Kai bus reikalinga tvarkyti ir metabolizuoti naują substratą, bus įjungtas genų rinkinys. Įjungus šių genų rinkinį, gaminami fermentai, kurie metabolizuoja naują substratą. Šis reiškinys vadinamas indukcija, o šio reiškinio sukeliančios molekulės (metabolitai) yra induktoriai.
Kita vertus, kai bakterijai reikalingas metabolitas yra tiekiamas iš išorės, bakterija slopina jo susidarymą, o tai stabdo tolesnį metabolito susidarymą bakterijoje. Šie inaktyvuoti genai yra vadinami reprezentatyviais, ir šis reiškinys yra žinomas kaip represijos arba grįžtamosios represijos.
Tačiau kai kurie genai yra nuolat ekspresuojami, kad būtų pasirūpinta normaliu ląstelių aktyvumu, tokiu kaip glikolizė. Šie genai yra žinomi kaip konstitutyvūs, o jų gaminami fermentai yra žinomi kaip konstituciniai fermentai, tokie kaip dehidrogenazės.
Operon koncepcija:
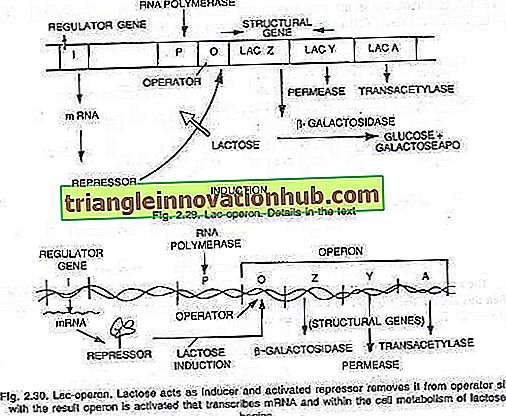
Francois Jacob ir Jacques Monod (1961), dirbdami Pasteur institute, Paryžiuje, pasiūlė, kad medžiagų apykaitos keliai būtų reguliuojami kaip vienetas. Jie ištyrė E. coli fermentų pobūdžio pasikeitimus, kai jie tiekiami su įvairių rūšių cukrumi. Jie nustatė, kad, pridėjus cukraus laktozę į E. coli kultūrą, jis sukelia tris fermentus, reikalingus laktozei suskaidyti į gliukozę ir galaktozę.
Šie naujai susintetinti fermentai yra - (i) p-galaktozidazė, (ii) permeazė ir (iii) transacetilazė. Jų teigimu, šių trijų fermentų sintezę kontroliuoja ilgas DNR segmentas, vadinamas operonu, kuris yra dalijamasi į operatoriaus svetainę O ir tris struktūrinius genus Z, Y ir A. Struktūrinių genų veikimą reguliuoja operatoriaus vieta su repressoriaus baltymo, gaunamo geno „i“, žinomo kaip reguliatoriaus genas, pagalba.
Genai yra išreikšti arba ne išreiškiami, priklausomai nuo to, ar įjungtas ar išjungtas operatoriaus jungiklis. Kai jungiklis yra įjungtas, trys genai RNR polimeraze transkribuoja į vieną RNS RNA, apimančią visus tris genus. Kiekvienas struktūrinis genas paprastai yra žinomas kaip cistronas, o transkribuotas ilgas mRNR, apimantis visus cistronus, yra žinomas kaip policistinis.
Operatoriaus jungiklio įjungimas arba išjungimas pasiekiamas naudojant baltymą, vadinamą repressoriumi. Kai šis baltymas susiejasi su operatoriumi (O) ir blokuoja jį, jungiklis išjungiamas ir trys genai (Z, Y, A) nėra išreikšti (žr. 2.28 pav.).
Lac-operonas:
Tai yra indukuojamo operono pavyzdys. Į ląstelę įdedama keletas laktozės molekulių, veikiančių fermento permeazės, kurios nedidelis kiekis randamas netgi esant represinėms sąlygoms. Po to šios kelios molekulės paverčiamos aktyvia laktozės forma, kuri jungiasi prie represoriaus (i geno produkto), todėl repressorius nebegali prisijungti prie operatoriaus jungiklio.
Kai operatorius neturi represoriaus, RNR polimerazė pradeda operono transkripciją, prijungdama promotoriaus vietą.
Šių trijų fermentų gamyboje prasideda laktozės metabolizmas. Fermentų sintezė tęsiama, išskyrus atvejus, kai ląstelėje vartojamos visos laktozės molekulės. Kai taip pat suvartojamos paskutinės laktozės molekulės, susietos su repressoriumi, neaktyvus repressorius tampa aktyvus ir prisijungia prie operatoriaus vietos (O), kad išjungtų operoną kaip įprasta (žr. 2.29 ir 2.30 pav.).
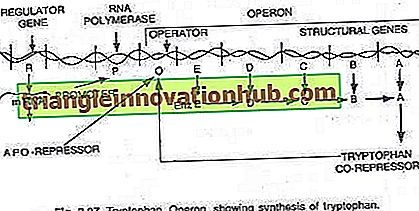
Tryptofano operonas:
Šiuo atveju operonas paprastai yra toks, kad transkripcija ir vertimas yra sintezuoti fermentus, reikalingus tam tikro metabolito sintezei ląstelėje. Tačiau operonas gali būti išjungtas, kai ląstelė nereikalauja metabolito arba metabolitas yra susidaręs pertekliumi. Tryptofano operonas susideda iš penkių genų (trp E, D, C, B ir A), koduojančių penkis fermentus (En z E, D, C, B ir A), kurie katalizuoja triptofano (aminorūgšties) sintezę ir todėl sudaro anabolinis kelias. Tryptofano buvimas padeda slopinti už jo gamybą atsakingų fermentų sintezę (žr. 2.27 ir 2.31 pav.).
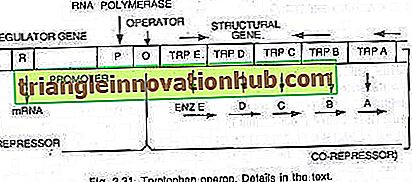
Šiuos penkis fermentus koduojančius penkis struktūrinius genus reguliuoja vienas operatoriaus jungiklis. Čia reguliuojamų genų gaminamas represoriaus baltymas pats nesusijęs su operatoriumi. Tačiau, kai pridedamas triptofanas, jis jungiasi prie represoriaus ir yra žinomas kaip represorius. Represoriaus-korepresoriaus kompleksas prisijungia prie operatoriaus ir blokuoja šio operono ekspresiją.
Tryptofano operono struktūra yra daugiau ar mažiau panaši į lac operono struktūrą, tačiau turi funkcinį variantą. Šiuo atveju R geno produktas yra lygiavertis lac operono genui. R genų produktas gamina baltymus, kurie savaime negali susieti su operatoriumi. Tai vadinama apo-repressoriumi.
Atliekant triptofaną, susidaro represorius, funkcinis represorius, kuris dabar jungiasi prie operatoriaus ir neleidžia operonui transkripuoti ir triptofano gamybai. Čia operono modelis paaiškina tiek indukcijos, tiek represijos reiškinį prokariotuose.

