„Golgi“ kompleksas: terminologija, atsiradimas, morfologija, struktūra ir kiti duomenys
Golgi kompleksas: terminologija, atsiradimas, morfologija, struktūra, struktūrinė ir biocheminė poliarizacija, cheminė sudėtis, funkcijos ir kilmė!
1898 m., Naudojant sidabro dažymo metodą, Golgi citoplazmoje atrado retikulinę struktūrą. Pavadinimas „Golgi aparatas“, paprastai suteiktas šiai struktūrai, yra painus, nes jis rodo aiškų ryšį su ląstelės fiziologiniais procesais.
Šiandien labiau tikslinga naudoti pavadinimą „Golgi medžiaga“ arba „Golgi kompleksas“, kad būtų nurodyta ši medžiaga, turinti ypatingų dažymo savybių. Kadangi jo lūžio rodiklis yra panašus į matricos, Golgi kompleksą sunku stebėti gyvose ląstelėse. Elektroninio mikroskopo naudojimas atskleidė aiškų šio komponento vaizdą ir atskleidė jo submikroskopinę struktūrą.
Jau daugelį metų „Golgi“ kompleksas buvo įvairių fiksavimo ir dažymo procedūrų artefaktas. Kitaip tariant, daugelis mokslininkų manė, kad daugelyje mikroskopinių procedūrų ir Golgi vadinamoji struktūra gyvoje ląstelėje iš tikrųjų neegzistavo. Guilliermondas (1923), Paratas (1927 m.), Walkeris ir A. Lien (1921), kilo abejonių dėl Golgi komplekso egzistavimo.
Golgi komplekso terminologija:
Holmgrenas nurodė Golgi kompleksą kaip trofosfį (Cajal jį pavadino Golgi-Holmgren kanalais). Bakeris vartojo terminą „lipochondria“ dėl numatomo lipidų kiekio. Terminas „Dalton Complex“ 1952 m. Buvo pateiktas po jos stebėtojo Daltono pavadinimo. Sjostrandas pasiūlė terminą cyto-membranos Golgi sistemai.
„Sosa“ pasiūlė šią „Golgi“ komplekso nomenklatūrą:
1. Golgiokinesis :
Golgi aparato padalinys branduolinio suskirstymo metu.
2. Golgiosomes :
Golgiogenezės gaminami korpusai vadinami Golgiosomomis, kurios yra apibūdintos kaip Golgi medžiaga bestuburiuose.
3. Golgiolizė :
Golgi aparato ištirpinimo procesas.
4. Gogiorrhexis :
Golgi aparato fragmentacija.
5. Golegiogenesis :
Golgi kūno formavimasis ir diferenciacija embriono vystymosi metu.
6. Golgio-cytoarchitecture :
Ląstelių struktūros tyrimas Golgi aparato atžvilgiu.
Įvykis:
Golgi kompleksas atsiranda visose ląstelėse, išskyrus prokariotines ląsteles (pvz., Mikoplazmą, bakterijas ir mėlynąsias dumblius) ir tam tikrų grybų, bryofitų ir pteridofitų spermos ląstelių, brandžių augalų sietų ląstelių ir brandžių spermos ir raudonųjų kraujo ląstelių. gyvūnų ląsteles.
Golgi komplekso morfologija:
Golgi komplekso morfologija priklauso nuo ląstelių, priklausomai nuo ląstelės, kurioje jie randami, tipą. Buvo stebimos dvi Golgi komplekso formos.

1. Lokalizuota forma :
Poliarizuotose stuburinių ląstelių (kurios turi bazę ir viršūnę), Golgi kompleksas atsiduria atskirai ir užima fiksuotą padėtį. Jis yra tarp branduolio ir sekretoriaus. Tai geriausiai matyti skydliaukės ląstelėse, kasos exokrininėse ląstelėse ir žarnyno ląstelėse.
2. Išskaidyta forma :
Kai kuriose specializuotose stuburinių (nervinių ląstelių ir kepenų ląstelių) ląstelėse, daugelyje augalų ląstelių ir bestuburių ląstelių, keletas Golgi komplekso vienetų yra išsklaidyti kartu su endoplazminio retikuliaus elementais. Kiekvienas vienetas yra vadinamas diktyvu. Kepenų ląstelėse vienam ląsteliui pasireiškia net 50 diktofosų, o tam tikrose augalų ląstelėse jų skaičius gali siekti iki šimtų.
Forma :
Golgi komplekso forma yra gana įvairi įvairiuose somatinių ląstelių tipuose. Net toje pačioje kameroje yra skirtingų funkcinių etapų skirtumai. Tačiau kiekvienos ląstelės tipas yra pastovus. Jis skiriasi nuo kompaktiškos masės iki išsklaidyto gijinio tinklo.
Skaičius:
Golgi stelažų skaičius vienoje ląstelėje labai skiriasi, priklausomai nuo ląstelių tipo - nuo kelių iki vieno. Kai kuriose ląstelėse yra vienas didelis, o Paramoeba atveju - du. Stereomyxa (Amoeba rūšis) yra daug Golgi kompleksų. Nervų ląstelėse, kepenų ląstelėse ir daugelyje augalų ląstelių taip pat yra keli Golgi kompleksai, kurių kepenų ląstelėse yra apie 50.
Dumblių rizoiduose yra daugiau nei 25 000 Golgi komplekso (Sievers 1965). Golgi kompleksas kai kuriose specializuotose ląstelėse netgi gali sudaryti didelę ląstelių tūrio dalį. Vienas iš pavyzdžių yra žarnyno epitelio taurė, kuri išskiria gleivius į žarnas; glikoproteinai gleivėse yra glikozilinti daugiausia Golgi komplekse.
Dydis:
Dydis taip pat yra kintamas. Jis yra didelis nervų ir liaukų ląstelėse ir mažas raumenų ląstelėse. Apskritai Golgi kompleksas yra gerai išvystytas, kol ląstelė yra aktyvioje būsenoje. Kai ląstelė sensta, kompleksas palaipsniui mažėja ir dingsta.
Pozicija:
Golgi komplekso padėtis yra santykinai nustatyta kiekvienam ląstelių tipui. Ektoderminės kilmės ląstelėse Golgi kompleksas yra poliarizuotas nuo embriono būklės tarp branduolio ir periferijos (Cajal, 1914). Sekretorinėse eksokrininėse ląstelėse, kurios paprastai turi tipišką poliarizaciją, Golgi kompleksas randamas tarp branduolio ir sekreto poliaus.
Endokrininėse liaukose šio organoido poliškumas yra kintamas, išskyrus skydliaukę, kur jis yra orientuotas į folikulo centrą. Jaunesnėse ląstelėse, dažniausiai senesnėse, dažniausiai jis yra vienoje branduolio pusėje, tačiau tam tikrais atvejais jis gali visiškai apsupti. Pelės ganglioninėse ląstelėse padėtis yra perinukle.
Išsami Golgi komplekso struktūra:
Daltonas ir Felixas (1954) apibūdino Golgi kompleksą žiurkių epididimyje po pirmųjų elektroninių mikroskopų.
Šis Golgi komplekso aprašymas yra sudėtinis, pagrįstas kelių autorių darbu:
1. Cisternae :
Cisternae arba saccules yra panašūs į lygų paviršių ER, ir sekcijoje jie rodomi kaip glaudžiai išdėstytų membranų atskirtų maišelių kaminai. Sakculių skaičius svyruoja nuo 4 iki 8 daugelyje gyvūnų ir augalų ląstelių tipų. Euglenoje šis skaičius gali siekti iki 20.
Kalkių membranos storis yra maždaug 60–70 °, apimantis apie 150 ° pločio ertmę, kurios kraštai dažnai išsiplėtę. Pasak daugelio autorių, yra du gerai apibrėžti cisternae veidai, ty išgaubti ir įgaubti; pastaroji paprastai vadinama subrendusiu ar formuojančiu ar distaliniu paviršiumi, o manoma, kad išgaubta pusė yra nesubrendusi arba susijaudinanti ar proksimalinė, o cisteranas yra lygiagrečioje matricoje ir yra atskirtas viena nuo kitos maždaug nuo 200 iki 300 A. 0 .
Tai, kas juos bendrai laiko, dar nežinoma, tačiau keliose ląstelėse yra tarp plonų elektronų nepermatomų sluoksnių, kartais tankių medžiagų, tarp tam tikrų regionų, kurie tam tikruose regionuose yra labiau pastebimi ir kuriems Amosas ir Grimstone (1968) taikė terminus mazgus. Mollenhauer ir kt. (1973) kai kuriais detalėmis tyrinėjo tarpkultūrinius elementus ir plokšteles tam tikrame Golgi komplekse.

2. Vamzdžiai :
Iš periferinės cisternae teritorijos kyla kompleksinis, anastomozinis plokščias 300–500 ° A skersmens vamzdelių tinklas. Clowes ir Juniper (1969) šį vamzdinį tinklą lygino su nėrinių disku.
3. Vesikėlės :
Pūslelės yra mažos lašelių formos maišeliai, kurie lieka prijungti prie tubulų cisternae periferijoje.
Jie yra dviejų rūšių:
a) lygios pūslelės :
Sklandžiai pūslelės yra nuo 20 iki 80 µm skersmens. Juose yra sekrecinė medžiaga (taip dažnai vadinama sekrecinėmis pūslelėmis), o tinklelio viduje esančios cisterninių vamzdžių galai yra nuplėšti. Dažnai yra daugiau nei viena vamzdelio jungtis ir, galbūt, užpildoma viena formuojanti pūslelinė.
b) Dengtos pūslelės :
Dengtos pūslelės yra sferinės iškyšos, kurių skersmuo yra apie 50 µm, o paviršius yra grubus. Jie randami organelio periferijoje, paprastai atskirų tubulų galuose ir yra morfologiškai gana skirtingi nuo sekrecinių pūslelių. Jų funkcija nežinoma.
4. Golgų vakuolai :
Tai yra dideli suapvalinti maišeliai, esantys Golgi brandinimo pusėje. Tai formuoja arba išplėstas cisternae, arba sujungiant sekrecines pūsleles. Vakuoliai pripildomi tam tikra amorfine arba granuliuota medžiaga.
Golgi kompleksas yra struktūriškai ir biochemiškai poliarizuotas:
„Golgi“ kompleksas turi du skirtingus veidus: cis, arba veido veidą ir trans, arba brandinimo veidą. Cis veidas yra glaudžiai susijęs su sklandžiu neapdorotos ER dalies. Sekretorinėse ląstelėse trans-veidas yra arčiausiai plazmos membranos artimiausias veidas: čia didelės sekrecinės pūslelės yra išskirtinai susijusios su Golgi kamino trans-veidu, o formuojančios sekrecinės pūslelės membrana dažnai yra nepertraukiama su paskutinio („trans-labiausiai“) cisternos trans-veidas.
Priešingai, mažos Golgi pūslelės yra tolygiau išdėstytos išilgai kamino, paprastai manoma, kad baltymai patenka į Golgi kaminą iš ER cis pusėje ir išeina iš kelių paskirties vietų trans pusėje; tačiau nėra žinomas nei jų tikslus kelias per Golgi kompleksą, nei kaip jie keliauja iš cisternos į cisterną išilgai kiekvienos kamino.
Du Golgi komplekso veidai yra biochemiškai skirtingi. Pavyzdžiui, tam tikrais atvejais, kai cis pusėje yra plonesni (ER panašūs) ir tie, kurie trans-pusėje yra storesni (plazminė membrana), galima nustatyti Golgi membranų storio kitimą.
Ryškesni yra rezultatai, gauti kai kuriems histocheminiams tyrimams kartu su elektronų mikroskopu, siekiant lokalizuoti tam tikrus baltymus Golgi komplekse. Kai kurie iš šių bandymų atskleidžia susijusią su membrana susijusią fermentą, kuris rodo, kad jų lokalizacija Golgi stekoje yra aiški.
Ypač intriguojantis biocheminis atradimas buvo atradimas, kad lizosominiai fermentai, tokie kaip rūgšties fosfatazė, yra koncentruojami trans-labiausiai colgos Golgi kamino ir kai kurių šalia esančių dengtų pūslelių. Tai rodo, kad šiame regione surenkami specifiniai lizosomoms paliekantys pūsleliai.
Sekretoriniai baltymai randami histocheminiais metodais visose sukrautose cisternaose, nors didelės sekrecinės pūslelės, kuriose šie produktai yra sukoncentruoti ir susieti tik su trans-dauguma Golgi cisterna.
Cheminė sudėtis:
Kalbant apie Golgi komplekso cheminę sudėtį, įrodyta, kad yra šių medžiagų:
1. Fosfolipidai :
Golgi membranų fosfolipidų sudėtis yra tarpinė tarp endoplazminių membranų ir plazmos membranų.
2. Baltymai ir fermentai :
Golgi kompleksas iš skirtingų augalų ir gyvūnų ląstelių rodo didelius baltymų ir fermentų kiekio skirtumus. Kai kurie fermentai yra ADPazė, ATPazė, NADPH citochromo-C reduktazė, glikozio 1 transferazės, galaktosilo 1 transferazė, tiamino pirofosfatas ir kt.
3. Angliavandeniai :
Augalų ir gyvūnų ląstelėse yra keletas bendrų angliavandenių komponentų, tokių kaip gliukozarinas, galaktozė, gliukozė, manozė ir fruktozė. Golgi augalui trūksta sialo rūgšties, bet žiurkės kepenyse ji būna didelė. Kai kurie angliavandeniai, tokie kaip ksilulozė ir arabinozė, yra tik augalų ląstelėse.
4. Vitaminas С :
Golgi komplekse saugomą vitamino С dalį parodė Tomitte. Pasak jo, „Golgi“ kompleksas saugo vitaminą С ir lėtai atpalaiduoja į citoplazmą, kad būtų išvengta ląstelių produktų oksidacijos.

„Golgi“ komplekso funkcijos:
1. Acrosome formavimasis spermiogenezės metu :
Spermos brendimo metu Golgi kompleksas atlieka svarbų vaidmenį formuojant akrosomą (Burgos ir Fawcett, 1955).

Ankstyvosiose stadijose Golgi pasirodo kaip sferinis kūnas, apimantis cisterną, išdėstytą lygiagrečiuose kaminuose ir daugelyje mažų pūslelių. Vėliau visuomet atsitraukė nuo cisterna. Vykstant vystymuisi, „Golgi“ kompleksas tampa netaisyklingas, o dideli vakuuminiai žiedai susidaro cisterninių maišelių išsiplėtimu.
Šių didžiųjų vakuolių centre arba vakuume yra tankus granulės, proakrosominės granulės. Ši granulė, kuri yra gauta iš Golgi komplekso, toliau auga vakuume pagal procesą, vadinamą įsiskverbimu. Ši vakuola ir granulė artėja prie branduolinės membranos priekinės poliaus, sudarančios akrosominę granulę.
Spermatido pailgėjimu, akrosominė pūslelinė plinta virš branduolinio paviršiaus ir pagaliau sugenda su branduoline membrana, sudarydama dangtelio medžiagą. Akrosominė granulė tampa akrosoma, kuri yra branduolio viršūnėje ir, matyt, apima tam tikrus fermentus, dalyvaujančius tręšimo procese.
2. Polisacharidų sintezė ir sekrecija:
Gobletų ląstelių tyrimai autoradiografijos ir elektroninės mikroskopijos būdu patvirtino sąveiką tarp baltymų sintezės, angliavandenių pridėjimo ir sulfacijos. Gyslos ląstelės iš gaubtinės žarnos gamina mucigen. Ši sekrecinė medžiaga turi daug angliavandenių.
Golgi kompleksas randamas tiesiai virš branduolio. Laisvo ląstelės paviršiaus link palaipsniui plečia mucigen granules. Golgi komplekso proksimalinė cisterna neturi jokio patinimo, bet tam tikru atstumu per stacką distalinė cisterna yra gana staiga konvertuojama į mucigen granules. Distalinė cisternae kas 2-4 minutes nuolat konvertuoja į mucigen granules. Kompensacijoje susidaro naujas proksimalinis cisterna.
3. Vaidmuo sekrecijoje:
Manoma, kad Golgi kompleksas atlieka tam tikrą vaidmenį ląstelių sekrecinėje funkcijoje. Tačiau kyla klausimas, ar jie pačios išskiria ar sintezuoja kai kurias medžiagas, arba tiesiog parduotuvės namai, kuriuose sekrecijos produktai, išskirti kitur ląstelėje, tiesiog saugomi ir koncentruojami.


Iš Palade et al. 1962 m. Šis sekrecijos ciklas dabar yra aiškiai apibrėžtas ir apima keturis etapus kasos acino ląstelių atveju ir jie yra:
(i) aminorūgščių įterpimas į baltymus apytiksliai endoplazminio tinklelio paviršiuje.
(ii) šių atsirandančių sekrecinių baltymų perkėlimas į neapdoroto endoplazminio retikuliaus cisternae.
(iii) šių baltymų pernešimas į Golgi kompleksą.
(iv) Zymogeninių granulių migracija į ląstelės viršūnę, į kurią jie išleidžiami į liumenus.
4. Golgi kūno vaidmuo oogenezėje :
Srivastava (1965) trumpai apžvelgė Golgi kompleksą oogenezės metu. Pasak Afzeliaus (1956), jūrų ežerų kiaušinio Golgi kompleksas, matomas elektronų mikroskopu, susideda iš plokščių maišelių sienelių, kurios kartais gali būti patinusios.
Yra keletas šių įstaigų skersinių padalijimų požymių. Sotelo (1959) ir Sotelo ir Porter (1959) aprašė Golgi kompleksą žiurkių kiaušialąstėse, kaip matėme elektronų mikroskopu, ir nustatė šio organelio branduolio lokalizaciją ankstyvosiose oocitose.
Kitame etape šie išsiskiria į fragmentus, o trečiajame etape šie žievės link. Visais šiais atvejais jų struktūra išlieka glaudžiai supakuotų plonų, dvigubų profilių (suplotų maišelių) ir sferinių pūslelių struktūra.
Ankstyvosiose oocitose kompleksas yra kompaktiškas. Vėlesnėse stadijose žievės zonoje yra išsklaidytų nedidelių profilių apsuptų nedidelių profilių paketų. Ankstyvosiose oocitose Golgi kompleksas ir centrosomas yra glaudžiai susiję.
5. Junginių absorbcija :
Hirsch ir kt. Nustatė, kad geležies cukrus tiekiamas gyvūnui, geležis absorbuojamas Golgi komplekse (Kedrowsky). „Van Teel“ parodė, kad „Golgi“ sistemos taip pat sugeria vario ir aukso junginius. Kedrowsky parodė, kad Golgi kompleksas Opalina gali absorbuoti bismutozę (albumino ir bismuto junginį) ir protargolį (albumino ir sidabro junginį). Taigi, Kirkmanas ir Severinghausas teigia, kad Golgi kompleksas veikia kaip kondensacijos membrana produktų koncentracijai į lašelius ar granules.
6. Augalų ląstelių sienelių susidarymas :
Augalų ląstelių sienelę sudaro fibriliai, kuriuose vyrauja polisacharidai ir kai kurie lipidai bei baltymai. Cytokinezės metu tarp dviejų dukterinių branduolių susidaro ląstelių plokštelė ir aplink ją yra membrana, kuri vėliau tampa dukterinių ląstelių plazmos membrana. Yra aiškių įrodymų, kad polisacharidai susidaro Golgi komplekse ir perkeliami į naują ląstelių sienelę, kuri yra nustatyta, kol ląstelės vis dar auga.

Golgi kompleksas taip pat prisideda prie medžiagų, tokių kaip pektinai ir hemiceliuliozės, kurios sudaro plazmos membranas atskiriančią ląstelių plokštelę.
7. Intracelulinių kristalų susidarymas :
Jūros izopode, Limnoria lingmorum, kuris yra burring forma, yra viduramžių, kurių ląstelės sudaro kristalai. Šie diapazonai yra iki 30 A ° ir 15 A ° storio. Įrodyta, kad šiuos kristalus sudaro Golgi kompleksas ir žinoma, kad juose yra baltymų ir geležies. Jie yra neuždengiančios membranos ir paprastai yra sferoidiniai. Jie rūpinasi sekrecijos veikla.
8. Pieno baltymų lašelių susidarymas :
Žindyvių žinduolių pelėse yra gaminami baltymų lašeliai, susiję su Golgi kompleksu. Šie lašeliai paprastai atsidaro ant ląstelės paviršiaus, suliejant jų uždengiančią membraną su plazmos membrana.
9. Lizosomų ir vakuolų susidarymas :
Pirminės lizosomos susidaro iš Golgi membranų taip pat, kaip sekrecinės pūslelės. Yra gerų įrodymų, kad diktofosmos sukaupia hidrolizinius fermentus savo brandesniuose regionuose. Nustatyta, kad kai kuriuose augalų ląstelėse esančiuose vakuoluose yra nedideli hidrolizinių fermentų kiekiai, ir manoma, kad jie buvo gauti iš Golgi komplekso.
10. Pigmentų susidarymas:
Daugelyje žinduolių navikų ir vėžio ląstelių Golgi kompleksas buvo apibūdintas kaip pigmentinių granulių (melanino) kilmės vieta.
11. Skysčių balanso reguliavimas :
Buvo pasiūlyta homologija tarp Golgi komplekso ir žemesniųjų Metazoa ir Protozoa kontraktinių vakuolių. Kontrakcinė vakuolė išleidžia iš perteklinio vandens iš ląstelės. Kai kuriuose protozojuose Golgi kompleksas taip pat susijęs su skysčių balanso reguliavimu.
Golgi komplekso kilmė:
Buvo pasiūlyti trys skirtingi šaltiniai, iš kurių gali kilti naujas „Golgi“ kompleksas:
1. Iš endoplazminio tinklelio :
„Essner“ ir „Novikоff“ (1962) ir „Beams“ ir „Kessel“ (1968 m.) Pasiūlė, kad Golgi cisternae kiltų iš ER. Neapdorotas endoplazminis retikulas po specifinių baltymų sintezės praranda ribosomas ir keičiasi į lygų ER. Mažos pereinamosios pūslelės sušvelninamos iš sklandaus ER. Jie migruoja į diktofosą.
Pasiekus formuojamą diktofosų veidą, šis saugiklis sudaro naują cisternę ir taip prisideda prie jo augimo. Sujungus šiuos pūslelius, formuojantis veidas nuolat susidaro naujas cisternae, o brandinimo metu senas cisternae suskaidomas į sekrecines vezikules. Taigi Golgi turi membraninio srauto reiškinį.
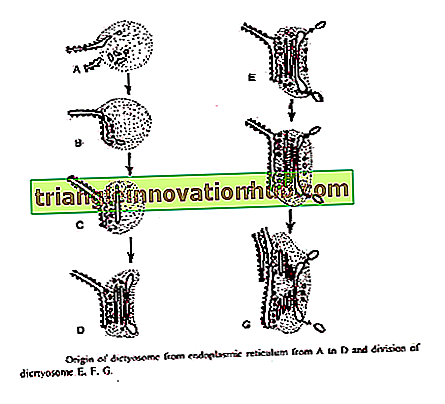

Bouchas (1965) apibūdino Golgi kilmę iš išorinės branduolinio voko membranos rudose dumblėse. Iš išorinės branduolinės membranos įsišaknijusios sėklidės, kurios sujungia cisterną formuojančiame diktofoso paviršiuje.
Atsiradimo zonų, susijusių su lygiagretaus ER arba branduoline membrana, buvimas, išskirtinių zonų atsiradimas aukštesniųjų augalų ramybės sėklose ir diktazomų susidarymas iš šių zonų dygsta sėklomis suteikia įrodymų, kad minėtos dvi teorijos yra susijusios su kilmės diktofosas.
3. Padalinus iš anksto egzistuojančius diktatus :
Pastebėta, kad ląstelių pasiskirstymo metu tiek augaluose, tiek gyvūnuose didėja diktofosomų skaičius ir kiekvienoje dukros ląstelėje diktofosų skaičius po pasidalijimo beveik lygus tėvų ląstelių skaičiui prieš padalijimą, iš skardinių ir kitų tiesioginių pastebėję dalijimosi ląsteles, buvo manoma, kad diktionos taip pat dalijasi ląstelių dalijimosi metu.

