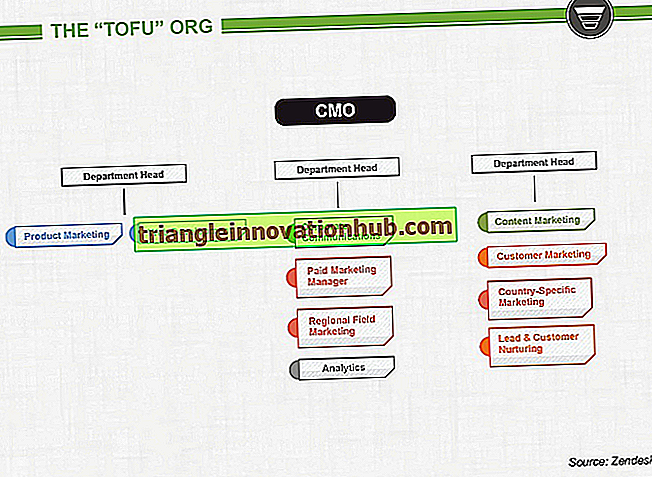
Trumpos užprogramuotos ląstelės mirties pastabos
Programuota ląstelių mirtis yra sukeltas ir surengtas procesas, kuriame ląstelė aktyviai dalyvauja kuriant savo mirtį.
Programuota ląstelių mirtis yra lemiamas veiksnys palaikant daugelio ląstelių tipų homeostatinį reguliavimą. Kiekvienai ląstelei yra laikas gyventi ir laikas mirti. Kiekvieną dieną iš kaulų čiulpų į apyvartą gaminami ir išleidžiami milijonai leukocitų. Leukocitų pusinės eliminacijos laikas yra tik kelias dienas. Po kelių dienų leukocitai miršta užprogramuota ląstelių mirtimi. Jei leukocitai nežūsta, bus kraujyje susikaupę leukocitai, kurie gali trukdyti kraujo skysčiui.
Nobelio prizas fiziologijoje ar medicinoje 2002 buvo apdovanotas trims mokslininkams - Sidnėjus Brenneriui, Robertui Horvitzui ir Johnui Sulstonui už atradimus, susijusius su „organų vystymosi ir programuotos ląstelės mirties genetiniu reguliavimu“. Jie naudojo nematodą „Caenorhabditis elegans“ kaip modelį ir nustatė pagrindinius genus, reguliuojančius užprogramuotą ląstelių mirtį. C. elegans yra mažas skaidrus kirminas, turintis trumpą kartos laiką, kuris padeda sekti ląstelių dalijimąsi tiesiai po mikroskopu. Programuota ląstelių mirtis arba apoptozė yra evoliuciškai konservuotas procesas. Stuburiniai gyvūnai sukūrė visas genų šeimas, panašias į nematodo ląstelių mirties genus.
Yra dvi priežastys, dėl kurių ląstelės gali užprogramuoti ląstelių mirtį arba apoptozę:
1. Norint tinkamai vystyti organizmą, reikalinga programuota ląstelių mirtis.
i. Apoptozė sukelia erškėčių uodegos rezorbciją jo metamorfozės metu į varlę.
ii. Ląstelių apoptinė mirtis veda prie audinių pašalinimo tarp vaisiaus pirštų ir pirštų.
iii. Endometriumo (vidinės gimdos gleivinės) atsitraukimas menstruacijų pradžioje vyksta apoptodis.
2. Programuojama ląstelių mirtis reikalinga šeimininkų ląstelių pašalinimui, kurios gali kelti grėsmę šeimininkui.
i. Virusai užsikrėtę ląstelės turi būti pašalintos iš kūno. Citotoksiniai T limfocitai (CTL) sukelia viruso infekuotų ląstelių apoptinį mirtį per fermentą granzyme.
ii. Po pageidaujamo ląstelių medijuojamo imuninio atsako periodo CTL turi būti pašalintas. Priešingu atveju CTL gali sukelti žalą kompiuteriui. CTLs sukelia apoptozę tarpusavyje. Be to, CTL gali sukelti apoptozę.
Ląstelės, kurioms užprogramuota ląstelių mirtis, turi daug morfologinių pokyčių, vadinamų apoptoze.
Ląstelėse, kurioms taikoma apoptotinė mirtis, stebimi šie morfologiniai pokyčiai:
i. Ląstelių tūrio sumažėjimas.
ii. Pakeitimai citozkelete, dėl to atsiranda membranos blebavimas.
iii. Chromatino kondensacija ir DNR skilimas mažais fragmentais.
Apoptinė ląstelė sulaiko daug membranų susietų apoptinių kūnų, kuriuose yra nepažeistų organelių. Apoptoziniai kūnai yra fagocituojami makrofagais. Apoptinės mirties metu dažymo ląstelės turinys nėra išleidžiamas į išorę (jei jos išsiskiria į išorę, jos paveiks netoliese esančias ląsteles ir sukelia uždegiminį atsaką). Apoptinės ląstelės turinys yra išleistas kaip membrana susietos apoptinės kūno dalys, o apoptiniai kūnai yra fagocituojami makrofaguose.
Ląstelės, kuri miršta dėl traumos (vadinamos nekrozė), savybės skiriasi nuo ląstelės, kuri miršta dėl apoptozės, savybių. Iš nekrotinės ląstelės išleistų proteazių gali pakenkti kaimyninės ląstelės, o iš nekrozinės ląstelės išsiskiriančios medžiagos stimuliuoja uždegiminį atsaką, dėl kurio uždegiminės ląstelės kviečiamos į nekrotinės ląstelės vietą. Kadangi ląstelinio turinio nutekėjimas iš apoptinės ląstelės yra kuo mažesnis, kad kaimyninės ląstelės nebūtų paveiktos, o uždegiminiai atsakai nesukelia.
Apoptozės stimulas ar signalas gali atsirasti ne iš ląstelių (išorinių), ar iš ląstelių (vidinės ar mitochondrijos).
i. Išoriniai signalai sukelia apoptozę (a) išorinės mirties, jungiančios ligandus prie ląstelių paviršiaus receptorių, prisijungimu ir (b) citotoksinių T ląstelių išskirto fermento granzimo poveikiu. Granzyme sukelia virusinių infekuotų ląstelių ir neoplastinių ląstelių apoptinę mirtį.
ii. Po įtampos ląstelėms (pvz., Spinduliuotės ar cheminių medžiagų ar virusinių infekcijų), prasideda vidiniai signalai. Apskritai, vidiniai signalai inicijuoja apoptozę, dalyvaujant mitochondrijoms.
Kaspazės:
Kaspazės yra citoplazminės proteazės. Kaspazės skiriasi nuo kitų proteazių tuo, kad kaspazės aktyviosiose vietose turi esminę cisteiną. Taip pat kaspazės išskiria tikslinius baltymus tam tikromis asparato liekanomis. Kaspazės yra citoplazmoje neveikliose prekursorių formose, vadinamose procaspazėmis. Procaspazės išskiriamos, kad būtų išlaisvintos aktyvios kaspazės. Citoplazmos procaspazes aktyvuoja daugelis ląstelių ir ląstelių ląstelių signalų. Iki šiol nustatyta daugiau kaip 10 žmogaus kaspazių.
Apoptozės metu chromosomų DNR yra išskirstoma į mažus nukleozominius vienetus. Citoplazminių fermentų kaspazės sukelia branduolinį skaidymą skirtingais mechanizmais.
i. Fermentas, aktyvuojantis DNR DNR (CAD) sukelia DNR fragmentaciją į nukleozominius vienetus (kaip matyti DNR kopėčių tyrimuose). Paprastai CAD egzistuoja kaip neaktyvus kompleksas su ICAD (CAD; DNS fragmentacijos faktorius 45). Kaspazės, įskaitant kaspazę-3, išsklaido ICAD ir atpalaiduoja CAD, o tai savo ruožtu sukelia greitą branduolinės DNR fragmentaciją.
ii. Polimero (ADP-ribozės) polimerazės (PARP) fermentas yra susijęs su sugadintos DNR taisymu. Kaspazė-3 išskiria PARP ir apsaugo nuo PARP DNR atstatymo funkcijų.
iii. Kaspazės inaktyvuoja branduolinio fermento DNR topoizomerazę II, kuri yra būtina DNR replikacijai ir remontui.
iv. Kaspazės suskaido branduolinės matricos ir citoskeleto struktūrinius baltymus ir lemia branduolio ir citoplazmos žlugimą. Laminai yra intranukleáris baltymai, palaikantys branduolio formą ir tarpininkauja tarp chromatino ir branduolinės membranos. Kaspazė-6 degraduoja laminus, dėl kurių susidaro chromatino kondensacija ir branduolio fragmentacija.
v. Kaspazės suskaido baltymus, reikalingus ląstelių ląstelių adhezijai, todėl apoptinės ląstelės išsiskiria iš gretimų ląstelių.
vi. Daugybė signalizuojančių molekulių, ląstelių ciklo reguliatorių ir transkripcijos faktorių skaidomi kaspazėmis.
Kaspazės veikia kaskados būdu, kaip ir komplemento komponentų kaskadinis aktyvavimas komplemento aktyvinimo metu. Yra keletas mechanizmų, kuriais galima aktyvuoti kaspazės kaskadą.
1. Mirtis sukelia ligandus prisijungia prie ląstelių paviršiaus receptorių.
↓
Iniciatoriaus kaspazės, tokios kaip kaspazė-8 arba kaspazė-10, yra aktyvuotos.
↓
Iniciatoriaus kaspazės kaskade aktyvuoja kitas kaspazes.
↓
Šis kaskadas galiausiai sukelia efektorinių kaspazių, tokių kaip kaspazės-3 ir kaspazės-6, aktyvaciją. Šios efektorinės kaspazės yra atsakingos už pagrindinių ląstelių baltymų skilimą, dėl kurių atsiranda morfologinių pokyčių ląstelėse, kurioms taikoma apoptozė.
2. Citotoksinės T ląstelės išskiria fermentus perforinus ir granzimą.
↓
Fermentas perforina smulkias skyles tikslinės ląstelės (pvz., Virusinės infekuotos ląstelės) ląstelių membranoje.
↓
Fermento fermentas patenka į ląstelę per poras ir aktyvuoja kaspazes 3, 7, 8 ir 10. Vėlesni įvykiai sukelia ląstelės branduolinę žalą.
3. Mitochondrija taip pat yra pagrindiniai kaspazės kaskados ir apoptozės reguliatoriai.
Iš mitochondrijų išsiskiriantis citochromas c jungiasi prie citopatinio baltymo Apaf-1.
↓
Pirmiau minėta sąveika yra stabilizuota ATP ir sukelia rato formos struktūrą, susidedančią iš 7 Apaf-1, citochromo c ir ATP molekulių. Ši ratų tipo struktūra, vadinama apoptosoma, leidžia į kompleksą įdarbinti 7 procaspazės-9 molekules.
↓
Procapazė-9 yra aktyvuota į kaspazę-9 (tikslaus procaspazės-9 aktyvavimo su kaspaze-9 mechanizmas nežinomas).
Bcl-2 baltymai ir apoptozė:
Bcl-2 genas (B ląstelių limfoma-2) koduoja baltymus, kurie slopina apoptozę. Iš pradžių Bcl-2 genas buvo aptiktas vėžio B ląstelėse, vadinamose B ląstelių limfomomis. Bcl-2 genas (B ląstelių limfomoje) yra chromosomų translokacijos lūžio taške. Bcl-2 genas perkeliamas į imunoglobulino sunkiosios grandinės lokusą, dėl kurio transkripcinis Bcl-2 geno aktyvinimas.
Todėl yra daugiau Bcl-2 baltymų gamybos. Padidėjęs Bcl-2 baltymų kiekis slopina normalius apoptotinių ląstelių signalus ir užkerta kelią apoptozei (manoma, kad Bcl-2 baltymų aukštojo lygio transformuotoje B ląstelėje padeda transformuotoms ląstelėms tapti vėžinėmis ląstelėmis, slopinant apoptinius signalus).
Apoptozę per mitochondrijų takus kontroliuoja baltymų šeima, susijusi su žmogaus onkoproteinu Bcl-2. Dauguma Bcl-2 baltymų yra integruoti citoplazminių organelių membraniniai baltymai, tokie kaip endoplazminis tinklas, išorinis branduolinis apvalkalas, vidinė plazmos membrana ir mitochondrija.
Įdomu tai, kad Bcl-2 šeimos baltymai suskirstomi į dvi grupes, turinčias biologiškai priešingą poveikį. Kai kurie baltymai skatina apoptozę (pvz., Bad ir Bax), o kiti (pvz., Bcl-2 ir BcIX L ) slopina apoptozę. Apoptinio signalo rezultatas ląstelėje gali priklausyti nuo pro-apoptinių ir anti-apoptinių Bcl-2 baltymų pusiausvyros. Pro-apoptinių baltymų perteklius gali sukelti ląstelių mirtį, o anti-apoptinių baltymų perteklius gali užkirsti kelią ląstelių mirčiai. Siūloma, kad Bcl-2 ir BclX L užkirstų kelią „Bax“ ir „Bad“ pro-apoptiniams veiksmams.
Mitochondrija ir apoptozė:
Mitochondrijos vaidina svarbų vaidmenį reguliuojant užprogramuotą ląstelių mirtį. Nežinomas mechanizmas, kuriuo Bax baltymas veikia apoptozėje.
Siūloma, kad Bax baltymai sudaro poras mitochondrijų išorinėje membranoje
↓
Mitochondrijų citochromo c ir apoptozės sukeliantis veiksnys (AIF) išeina per poras į citoplazmą.
↓
Citochromo c sudaro apoptosomą (su Apaf-1 ir ATP) ir aktyvuoja kaspazę-9.
Manoma, kad „Bcl-2“ ir „BcIXl“ užkerta kelią „Bat“ ir „Bax“ porų susidarymui mitochondrijoje.
Mirties receptoriai ir apoptozė:
Mirties receptoriai yra ląstelių paviršiuje esantys receptoriai. Sujungus su specifiniais ligandais, mirties receptoriai perduoda apoptinius signalus į ląstelę. Mirties receptoriai priklauso auglio nekrozės faktoriaus (TNF) genų super šeimai. Be apoptozės, mirties receptoriai taip pat sukelia daug kitų funkcijų.
Geriausiai žinomi mirties receptoriai yra:
i. Fas (CD95)
ii. TNFRl (TNF receptorius 1)
iii. TRAIL (su TNF susijęs apoptozės sukeliantis ligandas)
Apoptozė per Fas molekules:
Ląstelių paviršiuje yra transmembraninių mirties receptorių Fas molekulių. Fas citoplazminės dalys turi mirties sritis (DD).
↓
Citotoksiniai T limfocitai (CTL) savo paviršiuje išreiškia FasL. FasL (Fas ligandas) yra trimeris. Fas molekulių asociacija (tikslinių ląstelių paviršiuje) su FasL (ant CTL) skatina Fas molekulių trimerizaciją.
↓
Sumažinus Fas molekules, Fas klasterio ląstelių mirtys dominuoja kartu.
↓
Adapterinis baltymas, vadinamas FADD (su Fas susijusiu mirties domenu), susijęs su Fas mirties domenais.
↓
FADD turi domeną, vadinamą mirties efektoriaus domenu (DED). Mirties efektoriaus domenas leidžia prijungti procaspazes-8 (taip pat žinomas kaip FLICE) į CD95-FADD kompleksą [Baltymų CD95-FADD ir procaspazės-8 kompleksas žinomas kaip DISC (mirties sukeliantis signalizacijos kompleksas)].
↓
Procaspazė-8 yra skaldoma ir susidaro kaspazė-8. Kaspazė-8 sukelia vykdymo kaspazių, pvz., Kaspazės-9, veiklą.
„Fas“ molekulės vaidina svarbias „Apoptic“ funkcijas trijose situacijose:
1. Cytotoksinis virusų užkrėstų ląstelių žudymas tarpininkauja per Fas (viruso užkrėstų ląstelių) sąveiką su FASL ant CTL paviršiaus.
2. Aktyvuotų CTL paviršių Fas molekulės jungiasi su FasL (ant tos pačios ląstelės) ir veda prie CTL apoptozės. Aktyvintų CTL pašalinimas yra vienas iš mechanizmų, atsakingų už aktyvuotų CTL pašalinimą, kai šeimininkui nereikalaujama imuninio atsako.
3. Siūloma, kad ląstelės imuninės privilegijuotose vietose (pvz., Priekinėje akies kameroje ir sėklidėse) išreikštų aukštą FasL kiekį savo paviršiuje. Šių ląstelių FasL susieja su Fas molekulėmis uždegiminėse ląstelėse (kurios gali patekti į privilegijuotas vietas) ir sukelti uždegiminių ląstelių pašalinimą. Taigi, imuninės privilegijuotų vietų ląstelės išeina iš uždegiminių priepuolių ir išgyvena. Manoma, kad FasL molekulės ant ląstelių paviršiaus imuninės privilegijuotose vietose yra atsakingos už šių ląstelių privilegijuotą pobūdį.
Apoptozė per TNFR1:
Auglio nekrozės faktorius (TNF) gaminamas T ląstelių ir aktyvuotų makrofagų. TNF prisijungia prie TNF receptoriaus 1 (TNFRI) ir inicijuoja keletą poveikių ląstelėje.
i. TNF-TNFR1 surišimas gali sukelti NF-kB ir AP-1 aktyvaciją, o tai savo ruožtu sukelia daugybę priešuždegiminių ir imunomoduliuojančių genų.
ii. TNF-TNFR1 surišimas gali sukelti apoptozę.
TNF-TNFR1 surišimas lemia TNFR1 trimerizaciją ir TNFR1 ląstelių ląstelių mirties domenų klasterizaciją.
↓
Adapterio molekulė, vadinama TRADD (su TNFR susijęs mirties domenas), prisijungia prie TNFRI mirties srities.
↓
TRADD gali įdarbinti daug skirtingų baltymų į aktyvintą TNFRl1.
↓
TRAF2 (TNFR susieto faktoriaus 2) įdarbinimas sukelia NF-kB ir JNK / AF-1 signalizacijos kelio aktyvavimą.
1. FADD įdarbinimas sukelia apoptozės indukciją, įdarbinant ir praliejant procaspazę-8.
2. Kitas TNFRI junginys, vadinamas RAIDD, gali būti siejamas su kaspazės 2 įdarbinimu. Todėl atsiranda apoptozė.
TRAIL apoptozės indukcija (su TNF susijusia apoptozė, skatinanti ligandą):
TRAIL apoptozės indukcija daugeliu atvejų yra panaši į apoptozės indukciją per Fas. TRAIL jungiasi prie receptorių DR4 arba DR5 ląstelių paviršiuose ir sukelia apoptozę. Tačiau tikslūs mechanizmai dar nežinomi.
Yra baltymų, vadinamų sūnų baltymų (DCR1 ir CCR2), kurie konkuruoja su DR4 ir DR5 receptoriais, kad jie būtų pririšti prie TRAIL. Šie apgaulės receptoriai užkerta kelią apoptozei.
p53 ir apoptozė:
p53, DNR surišantis baltymas yra transkripcijos aktyvatorius. Kai DNR yra pažeista, pažeistos ląstelės padidina jų p53 baltymų gamybą. p53 yra stiprus apoptozės induktorius. Manoma, kad p53 veikia kaip DNR pažeidimo jutiklis ir yra vadinamas „genomo sergėtoju“.
Mutacijos p53 gene gali sutrikdyti ląstelės apoptinę mirtį. Daugelyje vėžio ląstelių tipų dažnai randamas mutuotas p53 genas, generuojantis defektinį p53 baltymą.
Virusai ir apoptozė:
Kai kurie virusai manipuliuoja apoptiniu mechanizmu savo naudai ir užtikrina jų išlikimą, užkertant kelią ląstelės, kurioje jie gyvena, apoptinei mirtis.
Vėžys ir apoptozė:
Siūloma, kad vėžio ląstelėse apoptotinis mechanizmas slopinamas. Todėl vėžinės ląstelės neišnyksta apoptozės būdu.
ŽIV ir apoptozė:
ŽIV infekuotiems asmenims palaipsniui mažėja CD4 + T ląstelių skaičius. CD4 + T ląstelių mažėjimo ŽIV infekuotiems asmenims mechanizmai nėra aiškiai žinomi. CD4 + T ląstelių apoptinė mirtis yra siūloma kaip vienas iš galimų mechanizmų. ŽIV užsikrėtę ir ŽIV neinfekuoti CD4 + T ląstelės ekspresuoja Fas molekules jų paviršiuje. ŽIV užsikrėtę CD4 + T ląstelės taip pat išreiškia aukštą FasL kiekį savo paviršiuje. ŽIV užsikrėtusių CD4 + T ląstelių FasL prisijungia prie Fas molekulių neužkrėstose CD4 + T ląstelėse ir sukelia apoptozę neužkrėstose ląstelėse, todėl sumažėja CD4 + T ląstelių skaičius. Manoma, kad ŽIV ne / genas yra atsakingas už aukštą FasL ekspresijos lygį ŽIV infekuotose CD4 + T ląstelėse.
Azoto oksidas ir apoptozė:
Azoto oksidas (NO) yra svarbi signalizavimo molekulė. NO dalyvauja reguliuojant įvairius procesus, pvz., Vazodilataciją, neuronų funkciją, uždegimą ir imuninę funkciją. NO gali sukelti apoptozę ir taip pat gali apsaugoti ląstelę nuo apoptozės. Įvairūs NO poveikiai priklauso nuo NO dozės, dalyvaujančių ląstelių ir daugelio kitų veiksnių.
Nustatyta, kad NO slopina leopocitų, hepatocitų, endotelio ląstelių ir trofoblastų apoptozę.
Yra daug mechanizmų, kuriais NO veikia kaip antisoptinis signalas:
i. Kaspazių, pvz., 1, 3 ir 8 kasazės, nitrozilinimas sukelia kaspazių inaktyvaciją.
ii. NO padidina šilumos šoko baltymą 70 ir dėl to blokuoja propaspazės-9 prijungimą prie apoptosomos.
iii. NEGALIMA Bcl-2 ir BcIX L, todėl citochromo c išsiskyrimas iš mitochondrijų yra slopinamas.